للحديث عن العشرين حمضًا أمينيًا التي تتكون منها الهياكل البروتينية والمعدلة ، سيكون من الضروري وصف ما لا يقل عن اثني عشر مسارًا استقلابيًا متخصصًا.
ولكن لماذا تستخدم الخلايا العديد من المسارات الأيضية التي تتطلب طاقة (على سبيل المثال لتجديد المواقع التحفيزية للأنزيمات) ، ولكل منها ميراث إنزيمي ، لتقويض الأحماض الأمينية؟ من جميع الأحماض الأمينية تقريبًا ، من الممكن الحصول ، من خلال مسارات متخصصة ، على المستقلبات التي تستخدم في جزء صغير منها لإنتاج الطاقة (على سبيل المثال ، من خلال استحداث السكر ومسار أجسام الكيتون) ولكنها ، قبل كل شيء ، تؤدي إلى تكوين مركبات معقدة جزيئات تحتوي على عدد كبير من ذرات الكربون (على سبيل المثال من فينيل ألانين وتيروزين ، يتم إنتاج الهرمونات في الغدد الكظرية المتخصصة لهذا الغرض) ؛ إذا كان من السهل إنتاج الطاقة من الأحماض الأمينية ، من ناحية أخرى ، فسيكون من الصعب بناء جزيئات معقدة تبدأ من الجزيئات الصغيرة: تقويض الأحماض الأمينية يسمح لها باستغلال هيكلها العظمي للحصول على أنواع أكبر.
يتم تحلل اثنين أو ثلاثة هيكتوجرام من الأحماض الأمينية يوميًا بواسطة فرد سليم: 60-100 جرام منها مشتق من البروتينات التي يتم إدخالها مع النظام الغذائي ولكن يتم الحصول على أكثر من 2 هيكتوجرام من معدل الدوران الطبيعي للبروتينات التي تشكل جزءًا لا يتجزأ من الكائن الحي ( يتم استبدال الأحماض الأمينية لهذه البروتينات ، التي تتلف بسبب عمليات الأكسدة والاختزال ، بأخرى ويتم تقويضها).
تعطي الأحماض الأمينية مساهمة في الطاقة من حيث ATP: بعد إزالة مجموعة α-amino ، يمكن للهيكل الكربوني المتبقي للأحماض الأمينية ، بعد التحولات المناسبة ، أن يدخل دورة كريبس. علاوة على ذلك ، عندما ينقص الإمداد بالمغذيات وتقل كمية الجلوكوز ، يتم تنشيط عملية استحداث السكر: يقال إن الأحماض الأمينية لتوليد الجلوكوز هي تلك التي يمكن إدخالها في عملية استحداث السكر بعد التعديلات المناسبة ؛ الأحماض الأمينية لتوليد الجلوكوز هي تلك التي يمكن تحويلها إلى البيروفات أو الفومارات (يمكن تحويل الفومارات إلى مالات الذي يترك الميتوكوندريون ، وفي السيتوبلازم ، يتحول إلى أوكسال أسيتات يمكن من خلاله الحصول على فوسفوينول بيروفات) .خل - أسيتات.
ما تم وصفه للتو هو جانب مهم للغاية لأن الأحماض الأمينية يمكن أن تعالج نقص السكر في حالة الصيام الفوري ؛ إذا استمر الصيام ، بعد يومين ، يتدخل استقلاب الدهون (لأنه لا يمكن مهاجمة هياكل البروتين كثيرًا) في هذه المرحلة ، نظرًا لأن تكوين الجلوكوز محدود للغاية ، يتم تحويل الأحماض الدهنية إلى أنزيم أسيتيل أ وجسم كيتون. من مزيد من الصيام ، يتكيف الدماغ أيضًا مع استخدام أجسام الكيتون.
يحدث نقل مجموعة α-amino من الأحماض الأمينية من خلال تفاعل التحويل ؛ الإنزيمات التي تحفز هذا التفاعل ، كما يقولون ، في الواقع ، الترانساميناسات (أو amino transferase). تستخدم هذه الإنزيمات عاملاً مساعدًا إنزيميًا يسمى فوسفات البيريدوكسال ، والذي يتدخل مع مجموعة الألدهيد الخاصة به. فوسفات البيريدوكسال هو نتاج فسفرة البيريدوكسين وهو فيتامين (ب 6) موجود بشكل رئيسي في الخضار.
Transaminases لها الخصائص التالية:
خصوصية عالية لزوج كيتوجلوتارات-غلوتامات ألفا ؛
تم تسميتهم على اسم الزوجين الثانيين.
تشتمل إنزيمات Transaminase دائمًا على زوج α ketoglutarate-glutamate ويتم تمييزها وفقًا للزوج الثاني المعني.
أمثلة:
L "ناقلة أمين الأسبارتات أي GOT (Glutamate-Ossal acetate Transaminase): ينقل الإنزيم مجموعة α-amino من الأسبارتات إلى α-ketoglutarate ، ويحصل على oxaloacetate و glutamate.
L "ترانس أميناز ألانين أي GTP (Glutamate-Pyruvate Transaminase): ينقل الإنزيم مجموعة α-amino من "ألانين إلى" α-ketoglutarate ، ويحصل على البيروفات والغلوتامات.
تستخدم الترانساميناسات المختلفة α-ketoglurate كمستقبل للمجموعة الأمينية من الأحماض الأمينية وتحويلها إلى جلوتامات ؛ بينما تستخدم الأحماض الأمينية المتكونة في مسار أجسام الكيتون.
يمكن أن يحدث هذا النوع من التفاعل في كلا الاتجاهين لأنهما ينكسران ويشكلان روابط بنفس محتوى الطاقة.
توجد الترانساميناسات في السيتوبلازم وفي الميتوكوندريا (وهي نشطة في الغالب في السيتوبلازم) وتختلف في نقطتها الكهربية.
إن الترانساميناسات قادرة أيضًا على نزع الكربوكسيل من الأحماض الأمينية.
يجب أن تكون هناك طريقة لتحويل الغلوتامات مرة أخرى إلى α-ketoglutarate: يتم ذلك عن طريق نزع الأمين.
هناك نازعة هيدروجين الجلوتامات إنه إنزيم قادر على تحويل الجلوتامات إلى كيتوجلوتارات ألفا وبالتالي تحويل المجموعات الأمينية من الأحماض الأمينية الموجودة في شكل جلوتامات إلى أمونيا. ما يحدث هو عملية الأكسدة والاختزال التي تمر عبر α-amino glutarate الوسيط: يتم إطلاق الأمونيا و α-ketoglutarate والعودة إلى الدورة الدموية.
بعد ذلك ، يمر التخلص من المجموعات الأمينية من الأحماض الأمينية عبر الترانساميناسات (التي تختلف وفقًا للركيزة) ونزعة الهيدروجين الجلوتامات ، والتي تحدد تكوين الأمونيا.
هناك نوعان من نازعة هيدروجين الجلوتامات: السيتوبلازم والميتوكوندريا. العامل المساعد ، وهو أيضًا الركيزة المشتركة لهذا الإنزيم هو NAD (P) +: يستخدم نازع هيدروجين الجلوتامات إما NAD + أو NADP + كمستقبل لتقليل الطاقة. يفضل الشكل السيتوبلازمي ، وإن لم يكن حصريًا ، NADP + بينما يفضل شكل الميتوكوندريا NAD +. الغرض من شكل الميتوكوندريا هو التخلص من المجموعات الأمينية: فهو يؤدي إلى تكوين الأمونيا (وهي ركيزة لإنزيم متخصص في الميتوكوندريا) و NADH (الذي يتم إرساله إلى السلسلة التنفسية). يعمل الشكل السيتوبلازمي في الاتجاه المعاكس أي أنه يستخدم الأمونيا و α-ketoglutarate لإعطاء الجلوتامات (التي لها وجهة تخليق حيوي): هذا التفاعل هو تخليق حيوي مختزل والعامل المساعد المستخدم هو NADPH.
يعمل نازع هيدروجين الغلوتامات عندما يكون من الضروري التخلص من المجموعات الأمينية من الأحماض الأمينية مثل الأمونيا (عن طريق البول) أو عندما تكون الهياكل العظمية للأحماض الأمينية ضرورية لإنتاج الطاقة: لذلك فإن هذا الإنزيم سيكون له عوامل تعديل سلبية الأنظمة التي تعتبر مؤشرًا توفر الطاقة الجيدة (ATP و GTP و NAD (P) H) وكمُعدِلات إيجابية ، الأنظمة التي تشير إلى الحاجة إلى الطاقة (AMP ، ADP ، الناتج المحلي الإجمالي ، NAD (P) + ، الأحماض الأمينية وهرمونات الغدة الدرقية).
الأحماض الأمينية (الليوسين بشكل أساسي) هي مُعدِّلات إيجابية لنزعة هيدروجين الجلوتامات: إذا كانت الأحماض الأمينية موجودة في السيتوبلازم ، فيمكن استخدامها لتخليق البروتين ، أو يجب التخلص منها لأنها لا يمكن أن تتراكم (وهذا ما يفسر سبب كون الأحماض الأمينية مُعدِّلات إيجابية ).
التخلص من الامونيا: دورة اليوريا
تتخلص الأسماك من الأمونيا عن طريق إدخالها في الماء من خلال الخياشيم ، وتحولها الطيور إلى حمض اليوريك (وهو منتج مكثف) وتزيله بالبراز. دعونا نرى ما يحدث في البشر: لقد قلنا أن نازع هيدروجين الغلوتامات يحول الغلوتامات في α- الكيتوجلوتارات والأمونيا لكننا لم نقل أن هذا يحدث فقط في الميتوكوندريا في الكبد.
تلعب ترانسامينازات الميتوكوندريا دورًا أساسيًا في التخلص من الأمونيا ، من خلال دورة اليوريا.

يتم تنشيط ثاني أكسيد الكربون ، على شكل أيون البيكربونات (HCO3-) ، بواسطة العامل المساعد للبيوتين المكون من كربوكسي البيوتين الذي يتفاعل مع الأمونيا ليعطي حمض الكرباميك ؛ يستخدم التفاعل التالي ATP لنقل الفوسفات إلى حمض الكرباميل مكونًا فوسفات الكرباميل و ADP (إن تحويل ATP إلى ADP هو القوة الدافعة للحصول على الكربوكسيبيوتين). يتم تحفيز هذه المرحلة بواسطة إنزيم فوسفات الكارباميل ويحدث في الميتوكوندريا. فوسفات الكرباميل والأورنيثين ركائز الإنزيم أورنيثين ترانس كارباميلاز مما يحولهم إلى سيترولين. يحدث هذا التفاعل في الميتوكوندريا (خلايا الكبد). يترك السيترولين المنتج الميتوكوندريا ، وفي السيتوبلازم ، يخضع لـ "تأثير"أرجينين سكسينات سينثيتاز: هناك اندماج بين الهيكل الكربوني للسيترولين والهيكل العظمي للأسبارتات من خلال هجوم محب للأنوية والتخلص اللاحق من الماء. يتطلب إنزيم سينثيتيز الأرجينين السكسينات جزيء ATP لذلك يوجد اقتران نشط: يحدث التحلل المائي لـ ATP إلى AMP والبيروفوسفات (يتم تحويل الأخير إلى جزيئين من أورثوفوسفات) عن طريق طرد جزيء d "الماء من الركيزة وليس بفعل ماء الوسط.
"الإنزيم التالي هو"أرجينين سكسيناز: هذا الإنزيم قادر على تقسيم سكسينات الأرجينين إلى أرجينين وفومارات داخل السيتوبلازم.
تكتمل دورة اليوريا بواسطة الإنزيم أرجيناز: يتم الحصول على اليوريا والأورنيثين. يتم التخلص من اليوريا عن طريق الكلى (البول) بينما يعود الأورنيثين إلى الميتوكوندريا ويستأنف الدورة.
تخضع دورة اليوريا لتعديل غير مباشر بواسطة الأرجينين: يشير تراكم الأرجينين إلى ضرورة تسريع دورة اليوريا ؛ يعد تعديل الأرجينين غير مباشر لأن الأرجينين يعدل بشكل إيجابي إنزيم الأسيتيل الجلوتامات المركب. هذا الأخير قادر على نقل مجموعة الأسيتيل على نيتروجين الجلوتامات: يتم تشكيل N-acetyl glutamate وهو مُعدِّل مباشر لأنزيم carbamyl-phospho synthetase.
يتراكم الأرجينين كمستقلب لدورة اليوريا إذا كان إنتاج فوسفات الكرباميل غير كافٍ للتخلص من الأورنيثين.
يتم إنتاج اليوريا فقط في الكبد ولكن هناك مواقع أخرى تحدث فيها التفاعلات الأولية.
يستخدم الدماغ والعضلات استراتيجيات خاصة للقضاء على المجموعات الأمينية. يستخدم الدماغ طريقة فعالة للغاية يستخدم فيها إنزيم تركيب الجلوتامين وإنزيم الجلوتاماز: الأول موجود في الخلايا العصبية ، والثاني موجود في الكبد. هذه الآلية فعالة للغاية لسببين:
يتم نقل مجموعتين من الأحماض الأمينية من الدماغ إلى الكبد في عربة واحدة ؛
الجلوتامين أقل سمية بكثير من الجلوتامات (يقوم الجلوتامات أيضًا بنقل الخلايا العصبية ويجب ألا يتجاوز التركيز الفسيولوجي).
في الأسماك ، تعمل آلية مماثلة على جلب المجموعة الأمينية من الأحماض الأمينية إلى الخياشيم.
من العضلات (الهيكلية والقلبية) ، تصل المجموعات الأمينية إلى الكبد من خلال دورة الجلوكوز ألانين ؛ الإنزيم المعني هو ترانس أميناز الجلوتامين-بيروفات: يسمح بتبديل المجموعات الأمينية (التي تكون في شكل غلوتامات) ، وتحويل البيروفات إلى ألانين ، وفي الوقت نفسه ، الجلوتامات إلى ألفا كيتوجلوتارات في العضلات ، وتحفيز عملية عكسية في الكبد.
تحتوي أيضًا Transaminases ذات المهام أو المواضع المختلفة على اختلافات هيكلية ويمكن تحديدها عن طريق الرحلان الكهربائي (لها نقاط متساوية كهربية مختلفة).
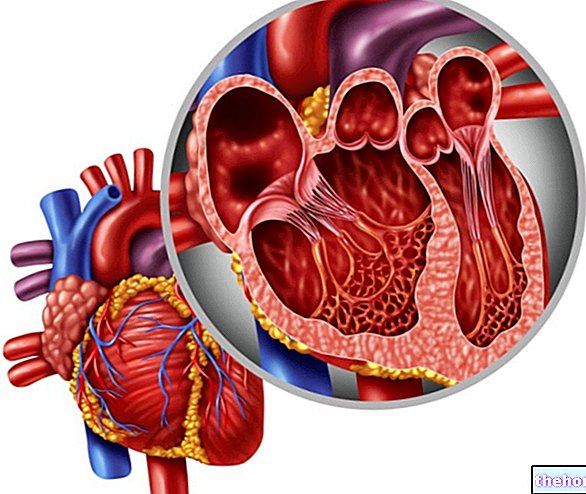
يمكن أن يكون وجود الترانساميناسات في الدم أحد أعراض تلف الكبد أو القلب (أي تلف الأنسجة في الكبد أو خلايا القلب) ؛ إن الترانساميناسات بتركيزات عالية جدًا في كل من الكبد والقلب: من خلال الرحلان الكهربائي ، من الممكن تحديد ما إذا كان الضرر قد حدث في خلايا الكبد أو القلب.



-ricotta-facile-e-veloce.jpg)