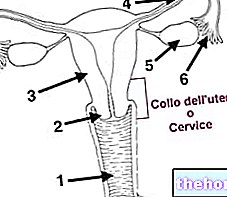
" الجزء الاول
الإنزيم الذي يحفز المرحلة الرابعة من دورة كريبس هو α-keto glutarate dehydrogenase؛ هذا الإنزيم عبارة عن مركب إنزيمي مشابه جدًا لبيروفات ديهيدروجينيز. كلاهما يتألف من 48-60 بروتينًا حيث يتم التعرف على ثلاثة أنشطة إنزيمية مختلفة ولها أيضًا نفس العوامل المساعدة الأنزيمية ؛ هي إنزيمات متشابهة جدًا لأنها تعمل على ركائز متشابهة: كلا من البيروفات و l "α- كيتو غلوتارات هي α- أحماض الكيتو. آلية عمل المجمعين الأنزيميين هي نفسها.
هجوم بيروفوسفات الثيامين على الكربونيل (C = O) من "α- كيتوجلوتارات ، يؤدي إلى نزع الكربوكسيل ويتكون مشتق بروبيل الكربوكسي هيدروكسي. مع النقل اللاحق إلى أميد الدهون ، تتم عملية الأكسدة والاختزال الداخلية ، والتي يتم من خلالها الحصول على مشتق كربوكسي أميد دهني أو سكسينيل ليبو أميد.
يتفاعل ليبو أميد السكسينيل مع الإنزيم المساعد A لإعطاء إنزيم السكسينيل A (الذي يستمر في دورة كريبس) وأاميد الدهون المختزل الذي يعاد أكسدة بواسطة FAD: يتم إعادة أكسدة FADH2 المتكون بواسطة NAD + ويتم الحصول على NADH. في هذه المرحلة ، تمت إزالة الكربون الثاني من الهيكل الكربوني ، على شكل ثاني أكسيد الكربون.
مجموعة الأسيل المرتبطة بالإنزيم المساعد A في شكل منشط ، أي أنها تحتوي على نسبة عالية من الطاقة: لذلك من الممكن استغلال طاقة إنزيم السكسينيل المساعد أ.

في المرحلة الخامسة من دورة كريبس ، يخضع أنزيم السكسينيل أ لفعل سكسينيل ثيوكيناز؛ تم وضع فرضيتين حول طريقة عملها: سنصف واحدة فقط من الاثنين لأنها الأكثر اعتمادًا.وفقًا لهذه الفرضية ، يتعرض الإنزيم المساعد للسكسينيل A للهجوم بواسطة النيتروجين الموجود في الهيستيدين (Hys) من الإنزيم: يتم تحرير الإنزيم المساعد A ويتم تكوين مقرب مشتق من الهيستيدين باعتباره وسيطًا ، وهو إنزيم السكسينيل (أو سوكسينيل-هيس). ) ؛ يعمل الأورثو فوسفات على هذا الوسط ، مما يؤدي إلى إطلاق السكسينات وتكوين الإنزيم الفوسفاتي ، وينتج الإنزيم الفوسفاتي ، الذي يهاجمه ثنائي فوسفات الغوانوزين (الناتج المحلي الإجمالي) ، غواسنوزين ثلاثي الفوسفات (GTP) ويتم إطلاق الإنزيم. من وجهة نظر الطاقة GTP = ATP: الرابطة التي توفر الطاقة هي نفسها في كلا النوعين (إنها رابطة أنهيدريد بين الفوسفوريل Β والفوسفوريل γ). في بعض الحالات ، يتم استخدام GTP كمواد ذات محتوى عالي من الطاقة ، ولكن عادةً ما يتم تحويل GTP إلى ATP من خلال عمل الإنزيم نيوكليوزيد ديفوسفو كيناز (NDPK) ؛ هو إنزيم موجود في الخلايا ويحفز التفاعل التالي:
N1TP + N2DP → N1DP + N2TP
عام NiTP ® نوكليوزيد ثلاثي الفوسفات
NiDP ® ثنائي فوسفات نيوكليوسيد عام
إنه رد فعل عكسي. في حالتنا يحدث:
GTP + ADP → الناتج المحلي الإجمالي + ATP
لذلك يمكن أن ينتقل إلى اليمين أو اليسار حتى بالنسبة للاختلافات الصغيرة في تركيزات الكواشف.
إذا استمرت دورة كريبس بهذه السرعة التي تؤدي إلى إنتاج ATP أعلى من متطلبات الطاقة ، فهناك نقص نادر في ADP بينما يوجد الكثير من ATP: التفاعل المحفز بواسطة nucleoside diphospho kinase هو ، إذن ، موجه إلى اليسار (يتراكم GTP إذا لم يكن للنيوكليوزيد diphospho kinase ركيزة كافية ، أي ADP). وبالتالي ، فإن GTP هي إشارة إلى توافر الطاقة وبالتالي تبطئ دورة كريبس.
تؤدي المرحلة السادسة من دورة كريبس إلى تكوين الفومارات بفعل نازعة هيدروجين السكسينات؛ يعطي هذا الإنزيم تفاعلًا خاصًا فراغيًا حيث يتم دائمًا تكوين غير المشبع (وهو ألكين) غير المشبع ، أي فومارات (بينما أيزومر رابطة الدول المستقلة هو ماليات). تم العثور على نازعة هيدروجين سكسينات على الغشاء الداخلي للميتوكوندريا ، بينما تنتشر جميع الإنزيمات الأخرى لدورة كريبس في جميع أنحاء الميتوكوندريا.
إنزيم نازعة هيدروجين السكسينات لديه FAD كعامل مساعد ؛ يتم تثبيطه بواسطة oxaloacetate (تثبيط التغذية المرتدة) بينما يحتوي على السكسينات والفومارات كمعدله الإيجابي (المنشط). دعنا نحاول فهم السبب ، من خلال القفز إلى المرحلة الأخيرة من دورة كريبس. المرحلة الأخيرة من تتطلب دورة كريبس طاقة ، لذا فإن الاحتمال الوحيد للحصول على أوكسالو أسيتات من المريض هو أن تركيز المريض مرتفع للغاية: المالات هو أحد المستقلبات ذات أعلى تركيز في الخلايا. حقيقة أن تركيز oxaloacetate يبقى منخفضًا عن طريق عمل سينسيز السترات. التفاعل المحفز بواسطة هيدروجيناز السكسينات هو ، إذن ، تفاعل ذاتي التغذية وهذه هي الطريقة الوحيدة لجعل تحويل مالات إلى أوكسالو أسيتات يحدث.
يجب أن يكون تركيز مالات الميتوكوندريا متوافقًا مع تركيز مالات السيتوبلازم: فقط عندما يكون تركيز مالات الميتوكوندريا مرتفعًا جدًا بحيث يضمن تحويل المالات إلى أوكسالو أسيتات (في دورة كريبس) ، يمكن أيضًا استخدام المالات في طرق أخرى (وهي السيتوبلازم): في السيتوبلازم يمكن تحويل المالات إلى oxaloacetate والتي يمكن الحصول على الأسبارتات من خلال عمل GOT (وهو ناقل أميناز) أو الجلوكوز من خلال تكوين الجلوكوز.
نعود إلى المرحلة السابعة من دورة كريبس حيث يتم تحفيزها بواسطة الإنزيم فوماراسي: يضاف الماء بطريقة الخصوصية الفراغية لصنع L-malate.
في المرحلة الأخيرة من دورة كريبس ، التي تحدثنا عنها بالفعل ، كان عمل نازعة هيدروجين مالات. يستخدم هذا الإنزيم جزيء NAD + لعمله التحفيزي.
بهذا نكون قد انتهينا من وصف المراحل المختلفة لدورة كريبس.
دورة كريبس قابلة للعكس تمامًا.
لزيادة سرعة دورة كريبس ، يمكن زيادة تركيز المستقلبات الموجودة في تلك الدورة ؛ تتمثل إحدى استراتيجيات زيادة سرعة دورة كريبس في تحويل جزء من البيروفات الذي يدخل الميتوكوندريا إلى أوكسال أسيتات (بفعل بيروفات كربوكسيلاز) وعدم تحويلها كلها إلى أنزيم أسيتيل أ: وبالتالي يزيد تركيز أوكسالو أسيتات الذي هو مستقلب دورة كريبس ، وبالتالي يزيد من سرعة الدورة بأكملها.
في دورة كريبس ، يتم تحويل ثلاث NAD + إلى ثلاثة NADH وواحد FAD إلى FADH2 ، علاوة على ذلك ، يتم الحصول على GTP: عن طريق توجيه الطاقة المختزلة التي تم الحصول عليها من دورة كريبس ، يتم إنتاج المزيد من ATP ؛ في السلسلة التنفسية ، يتم نقل القدرة المختزلة من NADH و FADH2 إلى الأكسجين: هذا النقل ناتج عن سلسلة من الإنزيمات الموجودة في غشاء الميتوكوندريا والتي تؤدي في عملها إلى إنتاج ATP.
عمليات السلسلة التنفسية هي عمليات مفرطة الطاقة وتستخدم الطاقة المحررة لإنتاج ATP ؛ والغرض من الخلية هو استغلال العمليات المفرطة للطاقة لجعل تخليق ATP يحدث. لكل جزيء من NADH يدخل في السلسلة التنفسية ، يتم الحصول على 2.5 جزيء من ATP ولكل FADH2 1.5 جزيء من ATP ؛ يرجع هذا التنوع إلى حقيقة أن FADH2 يدخل السلسلة التنفسية عند مستوى أقل من NADH.
مع القدرة المخفضة على التمثيل الغذائي الهوائي ، يتم الحصول على 30-32 ATP (219-233 كيلو كالوري / مول) بكفاءة تبلغ حوالي 33٪ (كفاءة التمثيل الغذائي اللاهوائي حوالي 2٪).