تعريف
الإنزيمات عبارة عن بروتينات يتم إنتاجها في الخلايا النباتية والحيوانية ، والتي تعمل كمحفزات لتسريع التفاعلات البيولوجية دون تعديل.
تعمل الإنزيمات من خلال الاندماج مع مادة معينة لتحويلها إلى مادة مختلفة ؛ يتم إعطاء الأمثلة الكلاسيكية عن طريق إنزيمات الجهاز الهضمي الموجودة في اللعاب والمعدة والبنكرياس والأمعاء الدقيقة ، والتي تؤدي وظيفة أساسية في الهضم وتساعد على تكسير الطعام إلى مكونات أساسية ، والتي يمكن بعد ذلك امتصاصها واستخدامها من قبل الجسم ، ومعالجتها بواسطة إنزيمات أخرى أو تفرز كنفايات.
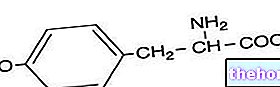
كل إنزيم له دور محدد: فالذي يكسر الدهون ، على سبيل المثال ، لا يعمل على البروتينات أو الكربوهيدرات. تعتبر الإنزيمات ضرورية لرفاهية الكائن الحي. ويمكن أن يتسبب النقص ، حتى في إنزيم واحد ، في حدوث اضطرابات خطيرة. ومن الأمثلة المعروفة على ذلك بيلة الفينيل كيتون (PKU) ، وهو مرض يتميز بعدم القدرة على استقلاب حمض أميني أساسي ، فينيل ألانين ، والذي يمكن أن يتسبب تراكمه في حدوث تشوهات جسدية وأمراض عقلية.
دراسة الكيمياء الحيوية
الإنزيمات هي بروتينات معينة لها خاصية كونها محفزات بيولوجية ، أي أن لديها القدرة على تحطيم طاقة التنشيط (Eatt) للتفاعل ، وتعديل مساره بحيث تكون العملية البطيئة حركيًا أسرع.
تزيد الإنزيمات من حركية التفاعلات المحتملة للديناميكا الحرارية ، وعلى عكس المحفزات ، فهي أكثر أو أقل تحديدًا: وبالتالي فهي تمتلك خصوصية الركيزة.
لا يشارك الإنزيم في القياس المتكافئ للتفاعل: لكي يحدث هذا ، يجب أن يكون الموقع التحفيزي النهائي مطابقًا لموقع البداية.
في الإجراء التحفيزي ، هناك دائمًا مرحلة بطيئة تحدد سرعة العملية.
عندما نتحدث عن الإنزيمات ، فليس من الصحيح الحديث عن تفاعلات التوازن ، فإننا نتحدث بدلاً من ذلك عن حالة مستقرة (الحالة التي يتم فيها تكوين مستقلب معين واستهلاكه باستمرار ، مع الحفاظ على تركيزه ثابتًا تقريبًا بمرور الوقت). عادة ما يكون ناتج التفاعل المحفز بواسطة إنزيم هو نفسه مادة متفاعلة لتفاعل لاحق ، يتم تحفيزه بواسطة إنزيم آخر ، وهكذا.
تتكون العمليات التي تحفزها الإنزيمات عادةً من متواليات من التفاعلات.
يمكن تلخيص تفاعل عام محفز بواسطة إنزيم (E) على النحو التالي:

E هو الانزيم
S هي الركيزة ؛
يمثل ES التقارب بين الإنزيم والركيزة ؛
P هو المنتج ؛
K هو ثابت معدل التفاعل.
يتحد الإنزيم العام (E) مع الركيزة (S) لتشكيل التقريب (ES) مع معدل ثابت K1 ؛ يمكن أن ينفصل مرة أخرى إلى E + S ، مع معدل ثابت K2 ، أو (إذا كان "يعيش" لفترة كافية ) يمكن أن ينتقل إلى تشكيل P بسرعة ثابتة K3.
يمكن للمنتج (P) ، بدوره ، إعادة الاتحاد مع الإنزيم وإصلاح التقريب باستخدام معدل ثابت K4.
عندما يتم خلط الإنزيم والركيزة ، هناك جزء من الوقت لم يحدث فيه الاجتماع بين النوعين بعد: أي ، هناك فترة زمنية قصيرة للغاية (والتي تعتمد على التفاعل) يكون فيها الإنزيم والركيزة لم يتم استيفائها بعد ؛ بعد هذه الفترة ، يتلامس الإنزيم والركيزة بكميات متزايدة ويتم تشكيل التقريب ES. في وقت لاحق ، يعمل الإنزيم على الركيزة ويتم إطلاق المنتج. ويمكن بعد ذلك القول إن c "هي فترة زمنية أولية لا يمكن فيها تحديد تركيز التقريب ES ؛ بعد هذه الفترة ، يُفترض أن الحالة المستقرة تم تحديده ، أي أن سرعة العمليات التي تؤدي إلى الحصول على المقربة تساوي سرعة العمليات التي تؤدي إلى تدمير المقربة.
ثابت Michaelis-Menten (KM) هو ثابت التوازن (المشار إليه في أول توازن موصوف أعلاه) ؛ يمكن القول ، مع تقريب جيد (لأنه يجب أيضًا مراعاة K3) ، يتم تمثيل KM من خلال النسبة بين الثوابت الحركية K2 و K1 (يشار إلى تدمير وتشكيل التقريب ES في التوازن الأول الموصوف أعلاه) .
من خلال ثابت Michaelis-Menten ، لدينا "مؤشر على التقارب بين الإنزيم والركيزة: إذا كان KM صغيرًا c" هو "تقارب كبير بين الإنزيم والركيزة ، فإن التقريب ES يكون مستقرًا.
تخضع الإنزيمات للتنظيم (أو التعديل).
في الماضي كان هناك حديث رئيسي عن التعديل السلبي ، أي تثبيط القدرات التحفيزية للإنزيم ، ولكن يمكن أيضًا أن يكون هناك تعديل إيجابي ، أي أن هناك أنواعًا قادرة على تعزيز القدرات التحفيزية للإنزيم.
هناك 4 أنواع من الموانع (تم الحصول عليها من التقديرات التي تم إجراؤها على نموذج لمطابقة البيانات التجريبية مع المعادلات الرياضية):
- تثبيط المنافسة
- تثبيط غير تنافسي
- تثبيط غير تنافسي
- تثبيط تنافسي
نتحدث عن التثبيط التنافسي عندما يكون الجزيء (المانع) قادرًا على منافسة الركيزة. بالنسبة للتشابه الهيكلي ، يمكن للمثبط أن يتفاعل بدلاً من الركيزة ؛ ومن هنا جاء مصطلح "التثبيط التنافسي" ، وتعتمد احتمالية ارتباط الإنزيم بالمثبط أو الركيزة على تركيز كليهما وعلى ارتباطهما بالإنزيم ؛ وبالتالي فإن معدل التفاعل يعتمد على هذه العوامل.
للحصول على نفس معدل التفاعل بدون وجود المثبط ، من الضروري الحصول على تركيز أعلى من الركيزة.
ثبت تجريبيا أنه في وجود مثبط ، يزيد ثابت Michaelis-Menten.
فيما يتعلق ، بدلاً من ذلك ، "التثبيط غير التنافسي ، والتفاعل بين الجزيء الذي يجب أن يعمل كمُعدِّل (مثبط إيجابي أو سلبي) و" الإنزيم ، يحدث في موقع يختلف عن الموقع الذي يكون فيه التفاعل يحدث بين الانزيم والركيزة. لذلك نتحدث عن تعديل خيفي (من اليونانية allosteros → موقع آخر).
إذا ارتبط المانع بالإنزيم ، فيمكنه إحداث تغيير في بنية الإنزيم ، وبالتالي يمكن أن يقلل من الكفاءة التي ترتبط بها الركيزة بالإنزيم.
في هذا النوع من العمليات ، يظل ثابت Michaelis-Menten ثابتًا لأن هذه القيمة تعتمد على التوازن بين الإنزيم والركيزة ، وحتى في وجود مثبط ، لا تتغير هذه التوازن.
ظاهرة التثبيط غير التنافسي نادرة ؛ المانع غير التنافسي النموذجي هو مادة ترتبط بشكل عكسي بالوصلة ES مما يؤدي إلى ظهور ESI:

قد يكون التثبيط الناتج عن الركيزة الزائدة غير قادر على المنافسة في بعض الأحيان ، حيث يحدث هذا عندما يرتبط جزيء الركيزة الثاني بمركب ES ، مما يؤدي إلى ظهور مجمع ESS.
من ناحية أخرى ، يمكن للمثبِّط التنافسي أن يرتبط فقط بعقد إنزيم الركيزة كما في الحالة السابقة: يؤدي ارتباط الركيزة إلى الإنزيم الحر إلى إحداث تعديل توافقي يجعل الموقع متاحًا للمثبط.
ينخفض ثابت Michaelis Menten مع زيادة تركيز المثبط: على ما يبدو ، يزداد تقارب الإنزيم مع الركيزة.
سيرين بروتياز
هم عائلة من الإنزيمات التي ينتمي إليها الكيموتريبسين والتربسين.
الكيموتريبسين هو إنزيم مُحلل للبروتين ومحلل مائي يقطع يمين الأحماض الأمينية الكارهة للماء والعطرية.
منتج الجين الذي يرمز لكيموتريبسين غير نشط (يتم تنشيطه بأمر) ؛ يتم تمثيل الشكل غير النشط من الكيموتريبسين بسلسلة بولي ببتيد من 245 من الأحماض الأمينية. يحتوي الكيموتريبسين على شكل كروي بسبب خمسة جسور ثاني كبريتيد وتفاعلات ثانوية أخرى (كهرباء ، قوى فان دير فال ، روابط هيدروجينية ، إلخ).
يتم إنتاج الكيموتريبسين بواسطة خلايا الكيموز في البنكرياس حيث يتم احتوائه في أغشية خاصة ويتم طرده من خلال قناة البنكرياس إلى الأمعاء ، في وقت هضم الطعام: الكيموتريبسين هو في الواقع إنزيم هضمي. تخضع البروتينات والمغذيات التي نتناولها من خلال النظام الغذائي للهضم ليتم تقليلها إلى سلاسل أصغر وامتصاصها وتحويلها إلى طاقة (مثل الأميليز والبروتياز يحلل العناصر الغذائية إلى جلوكوز وأحماض أمينية تصل إلى الخلايا عبر الأوعية الدموية يصلون إلى الوريد البابي ومن هناك يتم نقلهم إلى الكبد حيث يخضعون لمزيد من العلاج).
يتم إنتاج الإنزيمات في شكل غير نشط ولا يتم تنشيطها إلا عندما تصل إلى "الموقع الذي يجب أن تعمل فيه" ؛ بمجرد الانتهاء من عملهم ، يتم إلغاء تنشيطهم. لا يمكن إعادة تنشيط الإنزيم ، بمجرد تعطيله: للحصول على "عمل تحفيزي إضافي ، يجب استبداله" بجزيء إنزيم آخر. إذا تم إنتاج الكيميتريبسين في شكل نشط بالفعل في البنكرياس ، فإنه يهاجم الأخير: التهاب البنكرياس هو أمراض ناتجة عن إنزيمات الجهاز الهضمي التي يتم تنشيطها بالفعل في البنكرياس (وليس في المواقع المطلوبة) ؛ بعضها إذا لم يتم علاجه في الوقت المناسب ، يؤدي الى الموت.
في الكيموتريبسين وفي جميع سيرين بروتياز ، يرجع الإجراء التحفيزي إلى وجود أنيون الكحول (-CH2O-) في السلسلة الجانبية للسيرين.
سيرين بروتياز يأخذ هذا الاسم على وجه التحديد لأن عملها التحفيزي يرجع إلى سيرين.
بمجرد أن يؤدي كل الإنزيم عمله ، قبل التمكن من إعادة العمل على الركيزة مرة أخرى ، يجب استعادته بالماء ؛ "إطلاق" السيرين بواسطة الماء هو أبطأ مرحلة في العملية ، وهي هذه المرحلة الذي يحدد سرعة الحفز.
يحدث الإجراء التحفيزي على مرحلتين:
- تكوين الأنيون بخصائص تحفيزية (كحول الأنيون) والهجوم اللاحق للنواة على الكربون الكربوني (C = O) مع انقسام رابطة الببتيد وتكوين الإستر ؛
- هجوم الماء مع استعادة المحفز (قادر على ممارسة عمله التحفيزي مرة أخرى).
يمكن أن تتكون الإنزيمات المختلفة التي تنتمي إلى عائلة سيرين بروتياز من أحماض أمينية مختلفة ، ولكن بالنسبة لها جميعًا ، يتم تمثيل الموقع التحفيزي بواسطة الأنيون الكحولي للسلسلة الجانبية للسيرين.
فصيلة فرعية من سيرين بروتياز هي من الإنزيمات المشاركة في التخثر (والتي تتكون من تحول البروتين ، من شكله غير النشط إلى شكل آخر نشط). تضمن هذه الإنزيمات أن يكون التخثر فعالًا قدر الإمكان ومحدودًا في المكان والزمان (يجب أن يحدث التخثر بسرعة ويجب أن يحدث فقط في محيط المنطقة المصابة). يتم تنشيط الإنزيمات المشاركة في التخثر في سلسلة (من تنشيط إنزيم واحد ، يتم الحصول على مليارات الإنزيمات: كل إنزيم منشط ، بدوره ينشط العديد من الإنزيمات الأخرى).
التخثر هو مرض ناتج عن خلل في إنزيمات التخثر: يحدث بسبب التنشيط ، دون ضرورة (لعدم وجود إصابة) ، للأنزيمات المستخدمة في التخثر.
توجد إنزيمات تنظيمية (تنظيمية) وإنزيمات مثبطة للأنزيمات الأخرى: بالتفاعل مع الأخيرة ، فإنها تنظم أو تمنع نشاطها ؛ حتى منتج الإنزيم يمكن أن يكون مثبطًا للإنزيم ، وهناك أيضًا إنزيمات تعمل بشكل أكبر ، وكلما زاد وجود الركيزة.
ليسوزيم
اكتشف لويجي باستير ، من خلال العطس في طبق بتري ، أن المخاط يوجد في المخاط إنزيم قادر على قتل البكتيريا: الليزوزيم؛ من اليونانية: ليزو = ما الحجم ؛ زيمو = إنزيم.
الليزوزيم قادر على تحطيم جدار الخلية للبكتيريا. تحتاج البكتيريا والكائنات أحادية الخلية بشكل عام إلى هياكل مقاومة ميكانيكيًا تحد من شكلها ؛ يوجد داخل البكتيريا ضغط تناضحي مرتفع للغاية بحيث تجذب الماء. سينفجر غشاء البلازما إذا لم يكن هناك جدار خلوي يعارض دخول الماء ويحد من حجم البكتيريا.
يتكون جدار الخلية من سلسلة عديد السكاريد حيث تتناوب جزيئات N-acetyl-glucosamine (NAG) وجزيئات N-acetyl-muramic acid (NAM) ؛ يتم كسر الرابطة بين NAG و NAM بواسطة التحلل المائي. تعمل مجموعة الكربوكسيل من NAM ، في جدار الخلية ، في رابطة ببتيدية مع حمض أميني.
بين السلاسل المختلفة ، تتكون الجسور من روابط زائفة الببتيد: يرجع التفرع إلى جزيء ليسين ؛ الهيكل ككل متفرع جدًا وهذا يمنحه ثباتًا عاليًا.
الليزوزيم مضاد حيوي (يقتل البكتيريا): يعمل عن طريق إحداث صدع في جدار البكتيريا. عندما ينكسر هذا الهيكل (المقاوم ميكانيكيًا) ، تسحب البكتيريا الماء حتى تنفجر. تمكن الليزوزيم من كسر رابطة الجلوكوزيد β-1،4 بين NAM و NAG.
يتم تمثيل الموقع التحفيزي لليزوزيم بواسطة أخدود يمتد على طول الإنزيم الذي يتم فيه إدخال سلسلة السكاريد: يتم وضع ست حلقات غلوكوزيدية من السلسلة في الأخدود.
في الموضع الثالث من الأخدود c "يوجد خنق: في هذا الموضع يمكن وضع NAG واحد فقط ، لأن NAM ، وهو ذو أبعاد أعلى ، لا يمكنه الدخول. يقع الموقع الحفاز الفعلي بين الموضعين الرابع والخامس: نظرًا لوجود NAG في الموضع الثالث ، سيحدث القطع بين NAM و NAG (وليس العكس) ؛ وبالتالي ، يكون القطع محددًا.
الرقم الهيدروجيني الأمثل لعمل الليزوزيم هو خمسة. في الموقع التحفيزي للإنزيم ، أي بين الوضعين الرابع والخامس ، توجد سلاسل جانبية لحمض الأسبارتيك وحمض الجلوتاميك.
درجة التنادد: يقيس القرابة (أي التشابه) بين تراكيب البروتين.
هناك علاقة قوية بين الليزوزيم واللاكتوز سينثيز.
يصنع اللاكتوز المركب اللاكتوز (وهو سكر الحليب الرئيسي): اللاكتوز هو جلاكتوزيل جلوكوزيد حيث c "عبارة عن رابطة β-1،4 غلوكوزيدية بين الجلاكتوز والجلوكوز.
لذلك ، يحفز إنزيم اللاكتوز المركب رد الفعل المعاكس لذلك الذي يحفزه الليزوزيم (والذي بدلاً من ذلك يقسم رابطة الجلوكوزيد β-1،4)
إن إنزيم اللاكتوز المركب هو ثنائي ، أي أنه يتكون من سلسلتين بروتينيتين ، إحداهما لها خصائص تحفيزية وقابلة للمقارنة مع الليزوزيم والأخرى هي وحدة فرعية تنظيمية.
أثناء الحمل ، يتم تصنيع البروتينات السكرية بواسطة خلايا الغدة الثديية عن طريق عمل galatosyl-tranferase (له "تماثل تسلسلي بنسبة 40٪ مع الليزوزيم): هذا الإنزيم قادر على نقل مجموعة galactosyl من بنية عالية الطاقة إلى بنية بروتين سكري. أثناء الحمل ، يتم إحداث تعبير عن الجين الذي يرمز لـ galactosisyl-transferase (هناك أيضًا تعبير عن جينات أخرى تعطي أيضًا منتجات أخرى): هناك زيادة في حجم الثدي لأنه يتم تنشيطه الغدة الثديية (غير النشطة سابقًا) التي يجب أن تنتج الحليب أثناء الولادة ، يتم إنتاج α-lactalbumin وهو بروتين تنظيمي: إنه قادر على تنظيم القدرة التحفيزية لـ galactosyl-transferase (عن طريق تمييز الركيزة). يتم تعديل Galactosyl-transferase بواسطة α-lactalalbumin وهو قادر على نقل الجالاكتوزيل إلى جزيء الجلوكوز: تشكيل رابطة β-1،4 glycosidic وإعطاء اللاكتوز (lactose synthetase).
ومن ثم ، فإن الجالاكتوز ترانسفيراز يحضر الغدة الثديية قبل الولادة وينتج الحليب بعد الولادة.
لإنتاج البروتينات السكرية ، يرتبط galactosyl transferase بـ galactosyl و NAG ؛ أثناء الولادة يرتبط الألبومين اللبني بجالاكتوزيل ترانسفيراز مما يجعل الأخير يتعرف على الجلوكوز ولم يعد NAG لإعطاء اللاكتوز.